Introduction
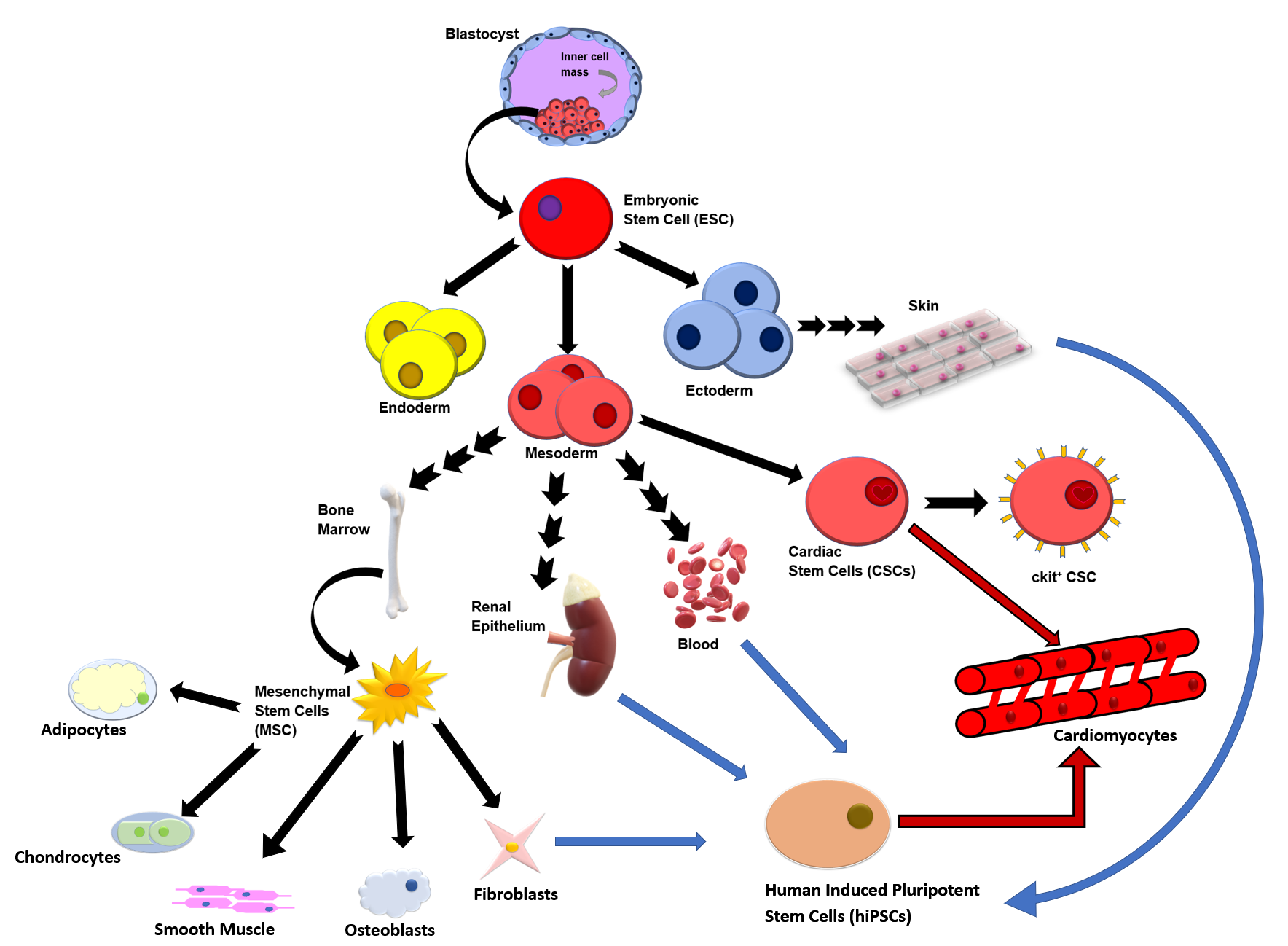
Despite advances in medicine, cardiovascular disease (CVD) remains the leading cause of death globally (for full list of abbreviations, see Table 1).1 CVD encompasses various conditions, including heart failure, atherosclerosis, coronary heart disease, myocardial infarction, arrhythmia, and heart valve dysfunction.1 Current non–stem cell therapies for CVD vary depending on the type and severity of the condition but usually include pharmacological interventions, surgical interventions, and lifestyle modifications in order to increase the functionality and protection of the heart.1 Existing therapies, such as coronary artery bypass graft surgery, protect against reperfusion and ischemic injury.1 However, such treatments do not address the seemingly irreversible remodeling of myocardial tissue, a process that occurs in response to chronic stress or injury, which typically leads to alterations in the cellular architecture of the heart.1 This process can impair the heart’s ability to pump blood effectively and lead to heart failure and other serious cardiovascular complications.1 Such complications are associated with frequent hospitalizations and overall poorer quality of life.1 While stem cells are not yet a standard treatment option for cardiovascular diseases (CVDs), the growing number of clinical trials exploring their use for various CVDs highlights their potential in regenerative medicine, including the future possibility of personalized tissue biomanufacturing.1,2 Stem cells include embryonic stem cells (ESCs), or cells derived from the inner layer of the blastocyst, and somatic stem cells, which can be induced further via nuclear content transfer or fusion with ESCs to create a diverse array of stem cells (Figure).1,3 For this review, we focused on chronicling existing research on mesenchymal stem cells (MSCs), induced pluripotent stem cells (iPSCs), and cardiac stem cells (CSCs) because these cells have already been used, without regard for microgravity, in clinical studies to improve treatment of heart pathology.

Improving the efficacy of stem cell treatments post cardiac injury is an object of future research, and budding evidence suggests that analyzing the effects of microgravity on stem cells could be a worthwhile endeavor.4 Microgravity conditions, like those simulated in clinorotation or environmental gravity in space, refer to environments with gravity less than that of Earth (1g = normal gravity).5 Clinorotation is a technique used in biological research in which organisms are slowly rotated to minimize the effects of gravity, theoretically simulating a microgravity environment to study how cells or tissues respond to the absence of gravitational forces.6 Although outside of the scope of this review, scaling up clinorotation for stem cell growth is possible, but it requires careful attention to factors such as consistent angular velocity and low sheer stress to maintain the integrity and effectiveness of the stem cells.7 The 3-dimensional nature of microgravity may give stem cells a beneficial environment not seen with the constant pull of Earth’s natural gravity, leading to a change in gene expression not seen on Earth.5 This difference increases myocardial differentiation in iPSCs, enriches transcription factors needed for cardiac metabolism, and increases expression of cardioprotective proteins such as Yes-associated protein isoform 1 (YAP1).4,8,9 Additionally, microgravity increases the expression of genes associated with improved cell survival and regeneration potency such as reelin (RELN) and cyclin D2 (CCND2).10 However, further examination is needed to determine the safety and efficacy of any derived treatment because evidence suggests that the mitochondria of cells cultured in microgravity do not regain normal gene expression on return to Earth; however, under simulated microgravity, the mitochondria of human iPSC cardiomyocytes show increased mitochondrial respiration compared with cultures under normal gravity.9,11 Microgravity also may have some unwanted effects on stem cells, such as upregulating hypertrophic pathways and increasing the size of the cells themselves.5,9 Moreover, the tumorigenic potential of stem cells must be considered, as well as the high level of radiation the cells are exposed to while onboard a spacecraft.5 This review will, therefore, explore the question: what are the positive and negative effects of microgravity on different stem cell lineages and the implications for the treatment of cardiovascular diseases?
Stem Cells in a Cardiovascular Lens
Regenerative medicine is an emerging field of inquiry, with stem cells as one of the primary focuses of research. While stem cell therapy requires more comprehensive clinical trials before US Food and Drug Administration approval for standard treatment of CVDs, the growing field of regenerative medicine provides an opportunity for its future clinical applications.1 ESCs and iPSCs display regenerative capabilities via their capacity to differentiate into cardiac cells.1 Methods to explore this regenerative ability for treatments analyzed in clinical studies include tissue patches, 3-dimensional tissue construct, stem cell–derived exosomes, and intracoronary, intravenous, and intramyocardial injection of stem cells (Table 2).1,12 Evidence from clinical trials suggests that stem cell therapies reduce the risk of future myocardial infarctions, have cardioprotective effects, and improve ejection fraction following myocardial infarction (Table 2).1
Three specific stem cell types of interest are bone marrow–derived MSCs, c-kit+ CSCs, and iPSCs (Figure). Transendocardial injections of bone marrow–derived MSCs have been shown to be effective in reducing infarct size and improving ejection fraction following induced myocardial infarction in mammalian models.16 Potential mechanisms for post–myocardial infarction cardiac repair and regeneration via bone marrow–derived MSCs include their ability for engraftment and differentiation, along with host-cell communication.16 Increasing evidence moves beyond the direct tissue regenerative aspect of stem cells to further highlight the communication of stem cells as they release paracrine factors, namely growth factors and cytokines, that seem to play a role in protection and recovery.1 These mediators appear to possess anti-inflammatory properties while also stunting fibrotic remodeling that causes irreversible damage and decreased function.1 The communication of MSCs appears to stimulate the recruitment and proliferation of CSCs, and their differentiation into cardiomyocytes, along with cardiomyocyte replication.16 This idea is supported by evidence that cardiac tissue under treatment of MSCs demonstrated significantly greater cardiomyocyte turnover compared with both the control and a concentrated conditioned medium from the MSC culture, representing MSC-secreted factors alone.16 Concentrated conditioned medium groups did not display the same improvements as the MSC-injected groups, suggesting that not only paracrine signaling, but also additional mechanisms play a distinct role in the ability of MSCs for cardiac repair.
Historically, the heart has been understood as a postmitotic organ. However, evidence for these c-kit+ CSCs opens a door for regenerative treatment of ischemic heart conditions.17 A direct comparison of the regenerative and cardioprotective effects of intramyocardial injected bone marrow–derived MSCs vs human c-kit+ CSCs shed light on the potential of this group of stem cells sourced directly from cardiac tissue.17 Compared with MSCs, human fetal CSCs displayed greater efficacy in attenuating left ventricular dilation and hypertrophy and infarcted scar size, while improving ejection fraction and left ventricular contractility. The CSCs also exhibited an ability for engraftment and cardiomyogenic differentiation.16,17 While fetal CSCs proved to be the most efficacious and are not a “clinically applicable” stem cell source, this limitation is mitigated by the results of the study’s comparison between fetal and adult human CSCs. Although adult CSCs are less potent, they still exhibit a potency equivalent to a 30-fold greater quantity of MSCs.17 Given the efficacy for the cardiac repair of MSCs and CSCs alone and the proposed communication of MSCs with CSCs, it is unsurprising that combined intramyocardial injection of human CSCs with human MSCs (hMSCs) displayed significantly greater scar size reduction and left ventricular diastolic and systolic function than either treatment alone or the placebo.18
Human iPSCs (hiPSCs) are also one of the primary and preferred sources for stem cell–derived cardiomyocytes in current research because they can be individualized and easily accessible compared with other sources, such as ESCs. Derived from host tissue, iPSCs are somatic cells that are genetically altered via viral transfection to behave similarly to ESCs, therefore gaining the capacity to differentiate into adult cells.1,3 Host tissue sources for iPSCs include, but are not limited to, blood, skin, renal epithelial cells, and fibroblasts (Figure).12 Additional attractive qualities of iPSCs include ion channels and signaling pathways identical to those of primary human cardiomyocytes, along with a capacity for prolonged culturing time.1 iPSCs may provide hope for those with CVD, but evidence suggests mixed efficacy in current applications.12 There is not only a lack of clarity on cell retention and survival due to hostile tissue microenvironments post myocardial infarction that promote cell death, but also an unclear relationship between cell dose and clinical efficacy.12
While promising, the use of stem cell therapy remains in clinical trials due to major limitations, including mixed results on efficacy and poor patient outcomes. Poor efficacy primarily derives from low cardiovascular retention due to stem cell death and washout following administration, along with potential immune rejection.1 Stem cell washout is the unintended loss or displacement of stem cells from their target site where they are intended to exercise their therapeutic effect.1 Poor patient outcomes in stem cell therapy of CVD include risk of chromosomal abnormalities, teratomas, cardiomyocyte loss post injury, deviation from target cardiac cell type, arrhythmia, and fibrosis.19 While adverse effects after administration of stem cells are relatively rare, the overwhelming number of potential adverse effects, along with uncertain efficacy, causes doubt for standard use within the next decade.1 While there are potential strategies that are currently being explored to address some of these limitations in cardiovascular stem cell therapy, one avenue that has yet to be adequately considered is the use of microgravity conditions on stem cells. Given the significance of MSCs, CSCs, and iPSCs to current stem cell research, improvements to these stem cell types via microgravity-induced mechanisms could incite major progress via increased efficacy and mitigation of major limitations.
Effects of Microgravity on Stem Cells
Emerging studies have delineated stem cell physiology within the context of microgravity as one potential and worthwhile approach to explore. Changes induced by microgravity involve cell signaling, gene expression, survival, differentiation, pluripotency, and morphology. Such changes have been reflected in both low-gravity environments, including satellites orbiting the Earth, and simulated-microgravity environments via clinorotation in laboratory settings. In space, low-gravity environments have a radiation component that 1g environments avoid.5 However, simulated microgravity is theoretical because it is inherently exposed to the effects of Earth’s gravity that is avoided in space.6 While microgravity’s full effect on stem cells is not yet known, the lack of gravity on the cell during growth and proliferation ultimately changes biochemical processes down the line.20
Stemness
Microgravity has been observed to increase the ability of stem cells to differentiate, proliferate, or self-renew into target adult cells, traits that fall under the umbrella term of stemness.20 Expression of nodal, mesoderm posterior protein 1 (MESP1), and NK2 homeobox 5 (NKX2-5), genes that act as early markers of CSCs and critical factors in heart development, are elevated in neonatal CSCs after simulated microgravity. MESP1 and NKX2-5 expression was increased by 29.32-fold and 15.05-fold, respectively.21 The MESP1-inducing gene Brachyury and MESP1-inducing protein octamer-binding protein 4 (Oct4) were also elevated by 3.6-fold and 5.5-fold, respectively, following simulated microgravity prior to differentiation.20–22 Furthermore, in mouse iPSCs tested during the Tianzhou-1 Space Mission, Oct4 demonstrated significantly quicker downregulation in space than on the ground on differentiation, indicating that cardiomyocyte formation was not only more robust in space but also accelerated.4
Pluripotency and Dedifferentiation
Microgravity affects multiple aspects of calcium signaling within stem cells, which likely plays a role in the ability of CSCs to maintain pluripotency.23 When exposed to microgravity via short-term simulated microgravity and International Space Station (ISS) culture, neonatal CSCs showed increases in protein kinase C α (PKCα) with increased protein kinase B (Akt) activation, both of which elevate calcium activity.23,24 Decreases in RhoA and CDC42 were also noted, with RhoA having major roles in actin cytoskeleton association, the Wnt planar cell polarity pathway, and modulation of intracellular calcium potentiation and oscillation.23,24 Experiments have shown that this biochemical change from decreased RhoA and CDC42 upregulates calcium pathways indicative of enhanced calcium handling.23,24 Increased calcium activity is associated with early cardiogenesis and maintenance of pluripotency.23,24 Thus, microgravity promotes the maintenance of pluripotency via calcium activation. Furthermore, WNT5A, a gene that is indicative of a more primitive cell state, also experienced elevated gene expression in short-term simulated microgravity conditions.23,24
Dedifferentiation is the necessary process that precedes cell cycle reentry and subsequent differentiation into the desired cell type, which in this case is specialized heart cells.21 Human adult and neonatal CSCs cultured onboard the ISS for 30 days expressed significantly higher levels of the dedifferentiation factors Sox2 and Klf4 compared with 1g controls.20 Given that Sox2 and Klf4 are factors that are used to reprogram differentiated somatic cells into iPSCs, their induction by microgravity may cause CSCs to develop into stem cells more similar to iPSCs.3 Notably, neonatal CSCs grown in microgravity exhibited downregulation of 2 microRNA (miRNA) molecules involved in cellular differentiation: miRNA-99a-5p and miRNA-100a-5p.21 The miR-99/100 families are highly conserved in mammals and have been studied in zebrafish, which possess endogenous cardiac regenerative mechanisms. Expression of miR-99/100 was high in zebrafish with uninjured hearts, but it became nearly undetectable on amputation of the ventricular apex and throughout subsequent endogenous cardiomyocyte regeneration.25 In this same study, knockout of miR-99/100 in zebrafish resulted in the upregulation of their target genes and dedifferentiation and proliferation of cardiomyocytes. Therefore, miR-99/100 is associated with maintaining a differentiated state in zebrafish, whereas ablating these miRNAs allows for dedifferentiation and regeneration. When observing mammalian heart regeneration, the inability to decrease heart injury–induced miR-99/100 elevation poses a hurdle for cardiac regeneration, and anti-miR administration has been hypothesized to overcome this.25 By downregulating miRNA-99a-5p and miRNA-100a-5p in human neonatal CSCs, microgravity may be a useful tool for facilitating cardiomyocyte dedifferentiation.21 Additionally, in vivo miR-99/100 downregulation in CSCs via mechanisms similar to those of zebrafish may be an exciting step forward in bypassing heart cell transplant limitations and reprogramming technologies in clinical settings.25
Cell Survival
Studies concerning cell signaling suggest that stem cell survival improves on exposure to microgravity environments via the amplification of certain factors and genes. An integral player in organ growth via cell proliferation that is affected by microgravity is the Hippo signaling pathway.8 Major factors of the Hippo signaling pathway include YAP1 and its downstream target SOD2. When induced, SOD2 prevents apoptosis by clearing reactive oxygen species.26 YAP1 and SOD2 levels were higher in human adult CSCs cultured in both real and simulated microgravity environments than in 1g.8 This is especially promising because YAP1 has been shown to protect against myocardial ischemic injury, promote cardiomyocyte survival, and prevent cardiac dysfunction after myocardial infarction, although without reducing cardiac hypertrophy.27,28 YAP1 is also shown to promote cardiac regeneration.29 Therefore, YAP1 and SOD2 promotion in microgravity-cultured stem cells suggests increased cardiac regenerative effects and reactive oxygen species clearance.26,29
In addition to the increased expression of Hippo signaling pathway factors, exposure of CSCs to microgravity resulted in the upregulation of 3 relevant genes: RELN, ubiquitin protein ligase E3 component N-recognin 3 (UBR3), and CCND2.10 RELN upregulation is associated with improving cell survival after heart injury.10 UBR3 regulates proteins that play a role in reducing genomic instability via improvements in DNA damage repair, cell survival, and transcription regulation.10 Upregulated CCND2 is linked to G1/S regulation, which is associated with regeneration potency and increased survival in hiPSC cardiomyocytes.10,30 Moreover, microgravity-associated upregulation of genes associated with DNA repair and telomere maintenance, including RAD50, RAD23, and Tert, has been noted in neonatal CSCs.21
Additional aspects of improved cell survival have been noted in MSCs in particular. MSCs grown in microgravity conditions for 2 weeks displayed diminished chemokine and, therefore, immune cell recruitment to damaged cells.5 Therefore, MSCs in microgravity seem to exhibit more potent immunosuppressive capacity, which holds the promising potential to address limitations centered around host immune rejection post stem cell transplant.5,19 Furthermore, the genomic integrity of MSCs did not appear to be compromised, even in the presence of higher genome-damaging space radiation, after two weeks aboard the ISS.5
Morphology
Microgravity has been shown to cause multiple morphological changes to stem cells, including changes in size and shape.6,10 Microgravity cultures of human CSCs aboard the ISS showed increased width, increased length, and more defined myofibrillar structure compared with 1g cultures, indicating a positive enhancement in structural development.10 Such phenotypic changes could be explained by the fact that myocyte enhancer factor 2 (MEF2), MEF2D, and specificity protein 1 (Sp1) transcription factor motifs are upregulated in flight compared with cells on Earth.9 Additionally, the expression of class II histone deacetylases HDAC10 and HDAC4 was depressed in flight, while the class I histone deacetylase HDAC8 was elevated in flight.9 This is especially interesting considering that when associated with MEF2, class II HDACs repress hypertrophy, while class I HDACs induce hypertrophy.9 Therefore, microgravity-induced depression of class II HDACs and elevation of class I HDAC8, in conjunction with increased MEF2 motifs, suggests augmented expression of hypertrophic pathways, potentially limiting future clinical applications.9 Similar to the negative hypertrophic effects of HDAC8 and MEF2, Sp1 is associated with hypertrophic phenotypic regulation, while upregulated MEF2D is associated with stress-dependent cardiac remodeling.31
Further, when hMSCs were subject to clinorotation-induced simulated microgravity, cell areas transitioned from fully “spread” shapes to more rounded morphologies, which is hypothesized to reduce cell migration. In concert with decreased observed motility and exhibited retracted cell area, this potential correlation between rounded morphologies and decreased cell migration suggests that microgravity may increase the localization of hMSCs at the desired tissue repair sites.6 This could pose an encouraging solution to decreased cell retention from stem cell washout, which is one major limitation of stem cell therapy.1 However, more research is needed to assess the extent to which these morphological changes are maintained in postflight long-term 1g conditions.5 Additionally, although these results show promise, other ISS studies have observed no change in morphology. Therefore, more research is needed to not only confirm the positive morphological changes, but also investigate the induction of negative hypertrophic pathways further for future clinical use.5,9
Electrophysiology
Electronic pacemaker implantation is widely accepted as a successful treatment for conduction disturbances, such as arrhythmias, but poses physical barriers such as limited battery and lack of neurohormone response.32 Traditional pacemaker cell research uses viral vectors as a means to upregulate electrically supportive factors.32 However, the development of autologous stem cell–derived pacemaker cells may provide a more efficacious method for electrical rewiring and improve quality of life, and microgravity may facilitate that process.32
MSCs have been proposed as a potential gene delivery system to create cardiac pacemaker cells.31 When cultured on the ISS, rat bone marrow–derived MSCs overexpressed C-X-C motif chemokine receptor 4 (CXCR4), which is a critical receptor for the electrical coupling and contraction of cardiomyocytes compared with ground controls.5,19 Furthermore, evidence supports that hMSCs with upregulated HCN2 channels, which control the pacemaker activity of the sinoatrial node, can more effectively promote cell to cell communication and simultaneous firing.32 Given that rat MSCs overexpressed CXCR4 in microgravity and that hMSCs in 1g amplified HCN2 channel expression, it would be meaningful to analyze the effects of microgravity in hMSCs on these proteins.
While we previously discussed increased calcium activity as a promoter of pluripotency, evidence shows it to also induce sinoatrial nodal genes.23,24 This concept coincides with the observation that microgravity increases not only calcium signaling, but also the induction of sinoatrial nodal genes in neonatal CSCs.23,24 This insight may be useful for the targeted production of sinoatrial node cells for clinical use. Further research is needed to investigate not only the feasibility of engineering biological pacemakers from CSCs, but also the quality of their spontaneous action potentials.
Evaluating Tumorigenicity and Teratoma Formation
While emerging studies of the effects of microgravity on stem cells have delineated the value of exploring this approach to regenerative therapy, certain limitations that would need to be addressed are the potential increased risk of tumorigenesis and teratoma formation. Historical applications of stem cell therapies have rare but present instances of unwanted proliferation leading to tumor formation.19 Therefore, while increased proliferation and survival of stem cells grown in space may be a favorable method of generating the large numbers of stem cells needed for clinical applications, it could also exacerbate this already-present risk of tumorigenic potential in stem cell therapies.5,12,19 This is particularly concerning considering radiation in space is 10-fold higher than on Earth, and cancer stem cells have shown similar increases in stemness and YAP activity when exposed to simulated microgravity.5,12,33 However, it is worth noting that microgravity may potentially induce anti-carcinogenic mechanisms considering MSCs aboard the ISS for two weeks showed no signs of damage to the cell genome and no evidence of tumor formation.5 Furthermore, efficient cardiomyocyte differentiation is associated with a lower likelihood of teratoma formation.19 Given that efficient differentiation is observed in space, there may be a link between microgravity and decreased risk of teratoma formation that has yet to be examined.4 Additionally, delivery of microgravity-exposed stem cell exosomes, rather than whole stem cells, may be a method to bypass the risk of tumor formation; however, further investigation of microgravity-induced stem cell exosomes is needed.12 Thus, although microgravity-grown stem cells have been observed to undergo increased proliferation and cell survival, certain tumorigenic qualities seen on Earth may be mitigated by a microgravity environment. Further research is required to answer the question of whether the microgravity would indeed increase or attenuate tumorigenic qualities for future clinical use.
Conclusion
Heart disease causes disability for millions across the world, but current non–stem cell therapies often do not focus on irreversible myocardial tissue remodeling. While current non–stem cell therapies have cardioprotective effects, patients with CVD still experience poorer quality of life.1 Nonetheless, clinical trials on stem cell therapies are a leap toward regenerative medicine for treating CVD, as evidence has shown increased left ventricular contractility and scar size reduction.17,18 While these studies are encouraging for stem cell therapies in CVD treatment, there is still a risk of teratoma growth, arrhythmias, and deviations from the target cell type.19 HiPSCs, MSCs, and CSCs are possible avenues for cardiac repair, with combination trials of MSCs and CSCs demonstrating impressive scar size reduction and improved cardiac function.1,18
Current stem cell treatment modalities are a promising avenue for future regenerative care, but they have concerning shortcomings in the current non–stem cell therapies of CVD. Stem cells grown in a microgravity climate may exhibit biochemical characteristics different from those grown in Earth’s natural gravity, potentially those needed to promote cardiac repair.19 Neonatal CSCs in microgravity displayed elevated Mesp1 and Oct4, indicating elevated heart development markers and more robust cardiomyocyte formation.4,22 In space, stem cells exhibited elevated PKCɑ and Akt, along with the subsequent promotion of calcium handling, which provide hopeful clues in understanding the enhanced pluripotency of these cells.23,24 Moreover, time in microgravity downregulated miRNA families, an effect associated with dedifferentiation, similar to endogenous cardiac capabilities studied in zebrafish; this similarity leads to hypothesized in-vivo mammalian cardiac regeneration, which would be a step in developing treatments.21,25 Unlike on Earth, stem cells cultured in microgravity upregulated the Hippo signaling pathway, forming cells with increased protective factors against myocardial ischemic injury through increased YAP1 and SOD2 expression.8,27,28 Trials also support the claim that microgravity-cultured stem cells have improved cell survival and upregulation of DNA repair-associated genes.5,21 Microgravity is also associated with the overexpression of the electrical contraction receptor CXCR4, which has implications for the production of biological pacemakers.19 Not only do stem cells in microgravity display these enhanced effects and more, but they also exhibit increased cardiomyocyte proliferation and earlier cardiogenesis, which are useful tools in improving currently limited proliferative capacity for prospective clinical use.23–25
The profound changes induced by microgravity present microgravity-grown stem cell lines as potential candidates for future clinical use in treating CVD. A common issue in stem cell research, and its associated therapies, is reduced cell survival post implantation and washout.1 These limitations may be ameliorated by microgravity’s effect on reducing cell migration because they display morphologic changes supportive of stem cell adherence to the desired location and promote cell survival. This limitation is compounded by the risk of unwanted differentiation and teratoma formation, although the relative risk is low.1,19,31 Although the known effects of microgravity on stem cells are in their infancy, microgravity-induced enhancement of characteristics supportive of cardiac regenerative potential greater than their 1g counterparts proves this to be a worthwhile avenue for further exploration. These findings may not only lead to more effective stem cell treatments to improve the lives of millions of people who have CVD, but also pioneer the future of regenerative medicine.